El olfato es el más sensible de los sentidos, ya que unas cuantas moléculas –es decir, una mínima cantidad de materia– bastan para estimular una célula olfativa. Detectamos hasta diez mil olores, pero como las estructuras olfativas, al igual que el resto de nuestro cuerpo, se deterioran con la edad, los niños suelen distinguir más olores que los adultos.
Desde el punto de vista químico, el olor es una sensación, una noción de estímulo y percepción producida en el olfato por la interacción de una sustancia orgánica con los receptores olfativos de los seres vivientes. Dicha interacción depende en gran medida de la volatilidad de la sustancia, ya que es necesario que las moléculas de la sustancia olorosa pasen a una fase gaseosa para que puedan llegar a la nariz y así ser percibidos; de igual forma, se requiere que puedan atravesar las membranas de las células epiteliales de la nariz y llegar a los receptores que enviarán la señal al cerebro, indicando la sensación del olor. Un punto importante que deben cumplir las moléculas olorosas es tener un peso molecular bajo, aunque se ha generado cierta controversia en este punto puesto que hay moléculas, tales como los esteroides, que poseen olor aunsiendo de gran tamaño. Otro aspecto en relación con el tamaño de la molécula es su influencia sobre el mecanismo de la quimiorrecepción, pues parece ser que cier tas incapacidades para percibir aromas (anosmias) en los humanos aumentan directamente conforme al tamaño molecular, lo que puede deberse a que las moléculas muy grandes no tienen un buen acomodo en el sitio receptor.
Para que haya estimulación deben entrar pequeños flujos de aire de alta frecuencia.
El estímulo químico, debe ser volátil, ingresa con el aire que respiramos ---- luego tiene relación con el mucus nasal (disolviéndose en proteínas del mucus) ---- actúa en los cilios de los receptores. El receptor también es tele-receptor, porque monitorea a distancia el estímulo.
9.2 Transducción olfativa
La transducción de las señales odoríferas comienza cuando moléculas químicas, se unen a los receptores específicos de membrana de los cilios (proteínas de membrana), ya sea de forma directa o a través de proteínas en el moco llamadas proteínas fijadoras de sustancias odoríferas.
Esta asociación a los receptores activa una proteína G específica de la sustancia odorífera que activa por su parte una adenilato cliclasa, lo que conduce a la generación de AMPc. Uno de los blancos del AMPc es un canal selectivo de cationes que cuando se abre permite el influjo de Na+ y Ca2+, este influjo crea un potencial de membrana, dando lugar al impulso eléctrico con que transmiten la información las neuronas.
En los cilios comienza la transducción. Moléculas olorosas se “acoplan” a proteínas receptoras.Los axones de las células mitrales constituyen el tracto olfativo que comunica el bulbo olfativo con el córtex olfativo.
La textura rugosa de la lengua se debe a un promedio de 10,000 papilas gustativas encargadas de identificar los 4 sabores básicos, dulce, salado, agrio y amargo, y un quinto sabor más llamado umami (sabor producido por el glutamato). Las papilas gustativas en la punta de la lengua detectan el sabor dulce, las de los lados, lo salado y ácido, y las de la parte de atrás, lo amargo. El gusto es el sentido que permite percibir los sabores. Las substancias estimulan los receptores sensoriales que se encuentran en las papilas gustativas de la lengua y en la pared posterior de la faringe. Estos responden con distinta intensidad a las diversas sensaciones gustativas, de modo que el sabor dulce se percibe especialmente en la punta de la lengua. El salado en la parte anterior. El ácido en los bordes y el amargo en la zona posterior. Los estímulos desencadenados son recogidos por los nervios glosofaríngeo, facial y vago, que los conducen hasta el núcleo gustativo del bublo raquideo, para ser transmitidos a las áreas gustativas del cerebro donde las sensaciones se hacen conscientes.
8.2 Transducción ácida
Ác. Clorhídrico (H+) + receptor (canal de Na) --- H+ ---provoca la apertura de los canales de Ca++ --- entra Ca++ --- depolarización ---- salen las vesículas --- transmisión de la señal. Además los H+ inhiben al canal de K+ ---- estimulando la depolarización de la membrana.
8.3 Transducción salada
El sabor salado se debe al catión sodio. Las células gustativas sensibles a este ión poseen en sus membranas canales iónicos. Cuando los cationes sodio entran en contacto con la membrana celular atraviesan estos canales y entran en las células gustativas. La acumulación de estos iones en el interior de las células provoca una despolarización de sus membranas y se genera así el impulso nervioso. Las moléculas que provocan estímulos dulces, cuando entran en contacto con las membranas de células gustativas en las microvellosidades, se unen con receptores. Estos son proteínas encajadas en la membrana, que interaccionan con mayor o menos intensidad con las moléculas “dulces”. Como resultado de esta interacción se activa el receptor, lo que desencadena una cascada de efectos dentro de las células que, finalmente, acaban generando el impulso nervioso
8.4 Transducción amarga
Presenta 2 posibles mecanismos:
1.-Sustancia amarga --- inhiba a los canales de K+ -- depolarización de la mb. -- entra Ca++ --- salen las vesículas --- transmisión de la señal.
2.-Sustancia Amarga + receptor para amargo---vía proteína G --- estimule fosfolipasa C --- liberación de IP3 (2º mensajero)---sale Ca++ de los depósitos --- vesículas salen --- transmisión de la señal.
8.5 Transducción dulce
Transducción del sabor Dulce:
Sucrosa (estímulo) + receptor del sabor dulce asociado a proteína G (alfa-gudocina) ---- adenililciclasa ---ATP en AMPc --a través de una Proteína Kinasa A. --- depolarización de mb. por fosforilación de canales de Ca++ -- entra Ca++ --- vesículas sinápticas salen --- transmisión del impulso nervioso. ---- Apertura de canales de K+
El ojo humano evolucionó en respuesta a la luz emitida por el Sol. Es por esto que nuestros ojos son sensibles a los colores que abarcan del amarillo al verde. La luz visible es una de las formas como se desplaza la energía. Las ondas de luz son el resultado de vibraciones de campos eléctricos y magnéticos, y es por esto que son una forma de radiación electromagnética (EM). La luz visible es tan sólo uno de los muchos tipos de radiación EM, y ocupa un pequeño rango de la totalidad del espectro electromagnético .
Sin embargo, podemos percibir la luz directamente con nuestros ojos, y por la gran importancia que tiene para nosotros, elevamos la importancia de esta pequeña ventana en el espectro de rayos EM.
7.2 El ojo y las células fotoreceptoras
El ojo humano es una máquina increíble, una de sus misiones es el registro de la presencia de luz, procesarla y enviarla al cerebro, en otras palabras: traducen las vibraciones electromagnéticas de la luz en un determinado tipo de impulsos nerviosos que se transmiten al cerebro.
El ojo humano cuenta con dos tipos de cèlulas para captar luz (fotoreceptores) y reciben su nombre según sus formas: bastones y conos. Los bastones son alargados y se encuentran entre los conos, son altamente sensitivos a la luz (sin color). Los conos a su vez son menos sensitivos a la luz pero altamente receptivos para el color y se dividen en tres grupos: los sensibles al color azul, al verde y al rojo. Los bastones proveen información sobre el brillo sin color y los conos sobre colores y detalles.
VISIÓN AL COLOR
Del espectro grande extraemos y ampliamos el espectro visible y representamos, de forma gráfica, qué es el color, qué son los colores, cual es el resultado de la suma de todos los colores (el blanco = luz blanca, luz día) y qué es la ausencia de color, el negro (la ausencia de luz). Antes de pasar al punto siguiente, el de la percepción del color, quedémonos con una idea clara, con un punto de referencia específico que es fundamental para entender este grupo de lecciones que tratan de los mecanismos de funcionamiento de los colores en los tejidos. Ese fundamento al que nos referimos es que el color no existe en la materia, no busquemos pues el color como algo palpable. En el mundo físico existe materia y energía. En esa energía vibratoria visible llamada luz es donde está aquello que en nuestro sistema visual suscita la sensación de color.
el intercambio de sustancias se realiza a favor del gradiente, esto es, en el sentido de los potenciales decrecientes, el requerimiento de energía externo al sistema es nulo; si, en cambio, el transporte se hace en contra del gradiente, se requiere el aporte de energía, energía metabólica en nuestro caso. Para que suceda este flujo, puesto que el agua se desplaza de un lugar muy concentrado a uno muy diluido en disolvente (en cuanto a soluto, se da la situación opuesta), y, por ello, lo hace a favor de gradiente, no se requiere un aporte de energía externo.
La naturaleza de las membranas biológicas, especialmente la de sus lípidos, es anfipática, lo que se traduce en que forman una bicapa que alberga una parte interna hidrofóbica y una externa hidrofílica, permite que surja una posibilidad de transporte, ladifusión simple o difusión pasiva, que consiste en ladifusiónde sustancias a su través sin gasto de energía metabólica y sin ayuda de proteínas transportadoras. En el caso de que la sustancia a transportar posea una carga neta, difundirá no sólo en respuesta a un gradiente de concentración, sino también al potencial de membrana
La fotoquímica es el estudio de las transformaciones químicas provocadas o catalizadas por la emisión o absorción de luz visible o radiación ultravioleta. Una molécula en su estado fundamental (no excitada) puede absorber un quantum de energía lumínica, esto produce una transición electrónica y la molécula pasa a un estado de mayor energía o estado excitado. Una molécula excitada es más reactiva que una molécula en su estado fundamental.
El fenómeno fotoquímico precisa de fases principales:
1. Recepción de la energía luminosa 2. Reacción química propiamente dicha.
Según se opere con una sustancia única o con un sistema de varios cuerpos en presencia, se realizará, bien una descomposición de la sustancia en sus elementos (fotólisis), bien una combinación de varios cuerpos en uno solo (fotosíntesis).
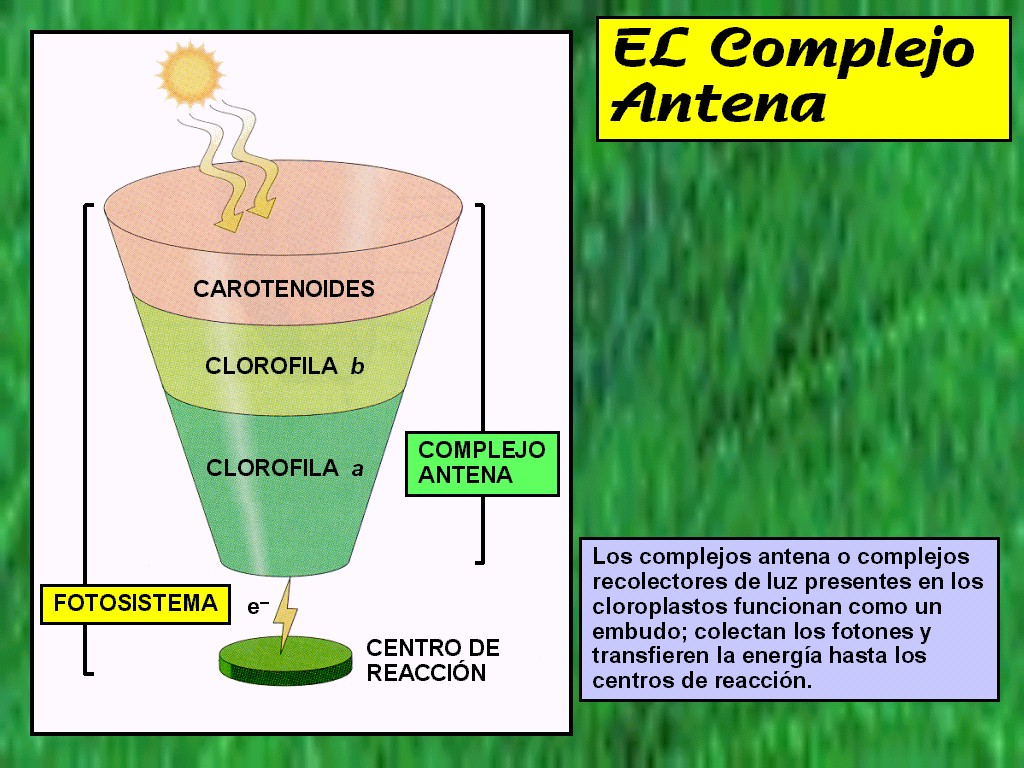
5.2 Pigmentos antena y captación de luz
los pigmentos antena sirven para captar la luz, no inician ninguna reacción en la mayoría de los casos. En las plantas fotosintéticas los principales pigmentos son la clorofila a y b, y las xantofilas en menor cantidad.
El pigmento capta la luz dependiendo de la longitud de onda de cada uno, después la energía en forma de fotones produce un cambio de conformación del pigmento, después se ioniza y se obtiene un electrón en forma de energía. Los pigmentos de los organismos fotosintéticos son la clorofila a y b, xantofilas, carotenoides, ficobilinas entre otros. Pero los pigmentos están presentes en todo el entorno, por ejemplo, el pigmento que nos permite ver a color es la rodopsina.
5.3 Cadena de transporte de electrones fotosintéticos
5.4 Análisis comparativo y evolutivo de la respiración y la fotosíntesis
La fotosíntesis y respiraciónson procesos íntimamente relacionados, producto de la fotosíntesis, que necesita de luz solar, dióxido de carbono y agua,liberando oxígeno a la atmósfera. La fotosíntesis es el mecanismo que tienen las plantas para producir energía química a partir de la luz del Sol, para su funcionamiento y para almacenamiento en forma de carbohidratos.
La respiración toma y consume este oxígeno liberando dióxido de carbono (y además agua). El oxígeno es necesario en los seres vivos aeróbicos para la respiración celular, y como un subproducto de este metabolismo de energía es que se desecha el CO2 (dióxido de carbono)
Ambos procesos son parte de un ciclo necesario para la vida de plantas y animales, donde se complementan mutuamente. Se puede decir que ambos procesos son parte de una relación dinámica, macroecológica.
Las
reacciones de óxido – reducción o REDOX son aquellas donde está involucrado un
cambio en el número de electrones asociado a un átomo determinado, cuando este
átomo o el compuesto del cual forma parte se transforma desde un estado inicial
a otro final.
La gran mayoría de las reacciones redox ocurren con liberación de energía.
La especie que suministra electrones es el agente reductor (especie que se
oxida) y la que los gana es el agente oxidante (especie que se reduce). Estos
hechos muestran que las reacciones redox se asemejan a las ácido-base según la
definición de Brönsted, pero en lugar de transferirse protones desde un ácido a
una base, en el caso de la oxido-reducción se transfieren electrones desde el
agente reductor al oxidante
3.2 Potencial redox
La capacidad oxidante es lo que se conoce como potencial de
oxidación (Eh), que cuanto más alto es, mayor es la capacidad oxidante del
sistema y mayor es la concentración de la forma reducida. El potencial de
oxidación se mide en voltios, aunque como su valor es muy pequeño se expresa
usualmente en milivoltios (mV).
El potencial redox es una medida de la actividad de los electrones. Está
relacionado con el pH y con el contenido de oxígeno. Es análogo al pH ya que el
pH mide la actividad de protones y el potencial redox mide la de los
electrones.
El potencial redox se calcula como:
Eh = 1, 234 - 0,058 pH + 0,0145 log (10) Po, siendo Po la presión parcial de
oxígeno expresada en atmósferas.
3.3 Radicales libres
los radicales libres son átomos o grupos de átomos que tienen
un electrón(e-) desapareado en capacidad de aparearse, por lo que son muy
reactivos. Estos radicales recorren nuestro organismo intentando robar un electrón de las
moléculas estables, con el fin de alcanzar su estabilidad electroquímica.
Una vez que el radical libre ha conseguido robar el electrón que necesita para
aparear su electrón libre, la molécula estable que se lo cede se convierte a su
vez en un radical libre, por quedar con un electrón desapareado, iniciándose
así una verdadera reacción en cadena que destruye nuestras células. La vida
biológica media del radical libre es de microsegundos; pero tiene la capacidad
de reaccionar con todo lo que esté a su alrededor provocando un gran daño a las
moléculas y a las membranas celulares.
Los radicales libres
producidos por el cuerpo para llevar a cabo determinadas funciones son
neutralizados fácilmente por nuestro propio sistema. Con este fin, nuestro
cuerpo produce unas enzimas (como la catalasa o la dismutasa) que son las
encargadas de neutralizarlos. Estas enzimas tienen la capacidad de desarmar los
radicales libres sin desestabilizar su propio estado
4.1 Hipótesis quimiosmótica y Potencial electroquímico de protón
Un gradiente electroquímico es una variación espacial tanto del potencial eléctrico como de la concentración de sustancia a través de una membrana. Ambos componentes son frecuentemente debidos a los gradientes iónicos (especialmente gradientes de protones), y de ellos puede resultar un tipo de energía potencial disponible para la realización de las distintas actividades celulares. Esto puede ser calculado como una medida termodinámica y combina los conceptos de energía almacenada en forma de potencial químico (que representa el gradiente de concentración de un ión a través de una membrana celular) y la energía electrostática, (lo que explica la tendencia de un ión a moverse en relación al potencial de membrana).
La magnitud termodinámica de interés en los procesos de transporte de un soluto a través de una membrana es el denominado Potencial Electroquímico del soluto. Se define como la variación de energía libre que ocurre cuando se transporta un mol del soluto a través de una membrana manteniendo constantes tanto las concentraciones del soluto en los compartimentos de llegada y de salida como el potencial de membrana. Es pues una magnitud que tiene sentido asumiendo condiciones de estado estacionario durante el proceso de transporte.
4.2 Estructura y función de la cadena respiratoria
Los electrones se transportan desde moléculas poco oxidantes hasta el oxígeno que es la molécula más oxidante de la cadena. Las moléculas que inician este transporte de electrones son NADH Y FADH, es decir son las moléculas menos oxidantes de la cadena. Una vez que los electrones son entregados al oxígeno, se forma Agua.
Debido a que la cadena sólo transporta electrones, los protones son bombeados hacia fuera de la mitocondria, lo que crea un gradiente de protones con una carga muy positiva afuera de la mitocondria y una carga muy negativa adentro. Este gradiente obliga a los protones a volver a entrar a la mitocondria y en el paso hacia adentro pasan por una enzima que forma un túnel de protones llamada ATP sintasa que con la fuerza de entrada de los protones, forma ATP.
4.3 Fosforilación oxidativa y síntesis de ATP
El fundamento de la fosforilación oxidativa es que la oxidación y la síntesis de ATP se acoplan mediante flujos de protones a través de la membrana. Ocurre en la membrana interna mitocondrial. Se realiza por medio de la ATP sintasa con sus dos complejos Fo y F1 formados por componentes móbiles y estáticos. Aprovechando el gradiente de protones que se crea en la cadena respiratoria la ATP sintasa sintetiza ATP a partir de ADP + Pi.
4.4 Genoma mitocondrial y enfermedades relacionadas
Las enfermedades mitocondriales son un grupo heterogéneo de alteraciones, caracterizadas por un fenotipo complejo en el que la mayoría de los pacientes presentan encefalopatía y pueden afectarse los músculos y otros órganos como corazón, hígado, riñones, retina, médula ósea, nervios periféricos y páncreas. La variación en sus manifestaciones clínicas puede explicarse no solo por la heterogeneidad en las mutaciones del ADNmt, sino también por mutaciones en el ADN nuclear que codifica gran parte de las subunidades proteicas de la cadena respiratoria, y más importante aún, por alteraciones del funcionamiento de las numerosas etapas que requiere esta cadena, las cuales están codificadas por ADN nuclear.
De todas ellas, la más frecuente es el síndrome de Leigh, que no se manifiesta hasta después del primer año de vida, cuando se produce una regresión con hipotonía y aparecen alteraciones del sistema nervioso central (síndrome piramidal, ataxia, oftalmoplejía, ptosis, nistagmo, distonía, temblor, atrofia óptica, neuropatía periférica y dificultad respiratoria).
Las mitocondriopatías asociadas a defectos de genes mitocondriales muestran gran variedad de fenotipos y se han descrito más de 70 mutaciones relacionadas con estas afecciones.
La bioenergética describe la transferencia y utilización de la energía en los sistemas biológicos. Utiliza las ideas básicas de la termodinámica, particularmente el concepto de energía libre. Los cambios en la energía libre (DG) proveen una cuantificación de la factibilidad energética de una reacción química y pueden proveer de una predicción de si la reacción podrá suceder o no.
La bioenergética se interesa sólo por los estados energéticos inicial y final de los componentes de una reacción, no del mecanismo o del tiempo necesarios para que el cambio químico se lleve a cabo. La bioenergética predice si un proceso es posible; la cinética cuantifica qué tan rápido ocurre la reacción.
Un estudio termodinámico se desarrolla sin necesidad de conocer la estructura molecular atómica de la materia y solo envuelve propiedades macroscópicas como presión, temperatura, volumen y las relaciones estre éstas.
La termodinámica es la ciencia que estudia los cambios
energéticos que acompañan los cambios físicos y químicos. .
Ley Cero de la Termodinámica
A esta ley se le llama de "equilibrio térmico". El equilibrio térmico
debe entenderse como el estado en el cual los sistemas equilibrados tienen la
misma temperatura.
Primera Ley de la Termodinámica
Esta primera ley, y la más importante de todas, también conocida como principio
de conservación de la energía, dice: "La energía no puede ser creada ni
destruida, sólo puede transformarse de un tipo de energía en otro".
Segunda Ley de la Termodinámica
La segunda ley dice que "solamente se puede realizar un trabajo mediante
el paso del calor de un cuerpo con mayor temperatura a uno que tiene menor
temperatura". Al respecto, siempre se observa que el calor pasa
espontáneamente de los cuerpos calientes a los fríos hasta quedar a la misma
temperatura.
Tercera Ley de la Termodinámica
El tercer principio de la termodinámica afirma que "el cero absoluto no
puede alcanzarse por ningún procedimiento que conste de un número finito de
pasos. Es posible acercarse indefinidamente al cero absoluto, pero nunca se
puede llegar a él"
La dirección y cantidad a la cual procede una reacción está determinada por el grado energía libre que dos factores cambian durante la reacción. Estos factores son la entalpía (DH, una medida del cambio de calor entre los reactivos y productos de la reacción) y la entropía (DS, una medida del cambio en el desorden de los reactivos y productos)
Ninguna de estas cantidades termodinámicas por si mismas es suficiente para determinar si una reacción podrá suceder espontáneamente en el orden en el que está escrita. Pero, cuando se combinan matemáticamente, es posible conocer la tercera, a partir del conocimiento de dos de ellas. La entropía no se puede determinar experimentalmente, se debe calcular a partir de G y H.
La mayor parte de procesos de transducción (conversión) de energía en la célula tienen lugar a través de membranas, conversión de energía redox (energía almacenada en electrones energéticos) en energía libre almacenada en un gradiente de potencial electroquímico durante la cadena respiratoriael uso de dicha
energía libre se utiliza para sintetizar ATP y para dar lugar al transporte de sustancias a través de membranas
Los seres vivos son máquinas químicas que utilizan la
energía química del ATP para hacer espontáneas a las reacciones endergónicas
Consumo de oxígeno = energía biológica
El 95 % del oxígeno consumido se consume en las mitocondrias
El 99 % del oxígeno consumido por las mitocondrias es reducido por la citocromo
oxidasa
El 100 % del oxígeno consumido por la citocromo oxidasa se utiliza para
sintetizar ATP por el mecanismo conocido como fosforilación oxidativa
2.1 El ATP como fuente de energía libre en los sistemas biológico
El trifosfato de adenosina o adenosín trifosfato (ATP-9 es un nucleótido fundamental en la obtención de energía celular. Está formado por una base nitrogenada (adenina) unida al carbono 1 de un azúcar de tipo pentosa, la ribosa, que en su carbono 5 tiene enlazados tresgrupos fosfato
Se produce durante la fotorespiración y la respiración celular, y es consumido por muchas enzimas en la catalisis de numerosos procesos químicos. Su fórmula es C10H16N5O13P3.
2.2 Estructura del ATP: inestabilidad, formación de híbridos de
resonancia y Potenciales de transferencia de fosfato
Las
reservas de ATP en el organismo no exceden de unos pocos segundos de consumo.
En principio, el ATP se produce de forma continua, pero cualquier proceso que
bloquee su producción provoca la muerte rápida (como es el caso de determinados
gases de combate diseñados para tal fin; o venenos como el cianuro, que
bloquean la cadena respiratoria; o el arsénico, que sustituye el fósforo y hace
que sean inutilizables las moléculas fosfóricas).
Las moléculas de creatina enlazan un fosfato mediante un enlace rico en energía
como el ATP. El ADP puede convertirse en ATP por acoplamiento con la hidrólisis
de fosfato de creatina. La creatina, por tanto, recicla el fosfato liberado por
la hidrólisis de la molécula de ATP original. Esto ayuda a mantener la energía
fácilmente movilizada sin agotar las reservas de ATP.
2.3 Hidrólisis de ATP y energía libre
Debido
a la presencia de enlaces ricos en energía (entre los grupos fosfato son los
enlaces anhídrido del ácido), esta molécula se utiliza en los seres vivos para
proporcionar la energía que se consume en las reacciones químicas. De hecho, la
reacción de hidrólisis de la adenosina trifosfato en adenosina difosfato y
fosfato es una reacción exergónica donde la variación de entalpía libre estándar
es igual a -30,5 kJ/mol:
La reacción de hidrólisis del ATP en adenosín monofosfato (y pirofosfato) es
una reacción exergónica donde la variación de entalpía libre estándar es igual
a -42 kJ/mo. La energía se almacena en los enlaces entre los grupos fosfato.Sin embargo, hay un nivel de entalpía a sobrepasar antes de liberar esta
energía (estado de transición). Esto explica por qué la hidrólisis de los
enlaces pirofosfato no sucede todo el tiempo. Las enzimas son capaces de
reducir ese umbral de entalpía para utilizar la energía liberada.
2.4 Hidrólisis de ATP acoplada a las reacciones bioquímicas no
espontáneas. NADH y FADH2 como fuentes de poder reductor
El metabolismo oxidativo de glúcidos, grasas y proteínas frecuentemente se
divide en tres etapas, de las cuales, el ciclo de Krebs supone la segunda. En
la primera etapa, los carbonos de estas macromoléculas dan lugar a moléculas de
acetil-CoA de dos carbonos, e incluye las vías catabólicas de aminoácidos (p.
ej. desaminación oxidativa), la beta oxidación de ácidos grasos y la
glucólisis. La tercera etapa es la fosforilación oxidativa, en la cual el poder
reductor (NADH y FADH2) generado se emplea para la síntesis de ATP